
BIOCHEMISTRY - DR. JAKUBOWSKI
This page has been compiled from several web pages and is not meant to be a cohesive chapter.
For our last class we will listen to a tape of a talk, Remembrance of Things Past, by Nobel Laureate Eric Kandel. He won the 2000 prize in Medicine and Physiology for his study of memory and learning, especially in the sea snail, Aplysia. The information in the following page comes from web links (references) and are threaded together to present the background to the talk we will hear on Monday. Please read this thoroughly so you can be prepared to get the most out of his talk.
Next Yr: Handout from talk
The are four different kinds of kinds of learning:
Kandel focuses on sensitization in his talk. All four types are summarized in this review.
http://www.brembs.net/learning/aplysia/
So what Aplysia behavior could one use to train it? The main focus on Aplysia neurobiology has been on classical conditioning of the defensive withdrawal reflex. A light tactile stimulus that would normally not produce any defensive reflex in the animal, is repeatedly paired with a strong noxious stimulus, usually an electric shock. After a few pairings, the animal shows the defensive reflex to the light tactile stimulus. Still, classical conditioning has been studied in a variety of animals, why Aplysia? The nice feature that makes Aplysia so attractive for neurobiologists is its large brain cells (neurons). The soma of one motor neuron can measure up to 1mm in diameter, which makes it relatively easy to study the physiology of these cells to find out how they accomplish learning.
For operant conditioning, on the other hand, one needs a spontaneous behavior that can be modified by reinforcement, so a reflex is not suitable. Well, about the only thing these animals do in their natural environment is to eat and reproduce. Naturally, their feeding behavior has received some attention from neurobiologists interested in operant conditioning (see videos of the Aplysia biting behaviour). From this research it was known that the esophageal nerve (En) had something to do with reinforcement. It is believed that sensory receptors in the buccal cavity transmit gustatory stimuli to the buccal ganglion via this nerve. We performed in vivo recordings from the anterior branch of En in feeding Aplysia via surgically implanted electrodes
from http://www.sahs.uth.tmc.edu/brainsurf/aplysia2.html

Sensory neurons are special nerve cells that can change the energy of a touch into chemical and electrical signals which are used to communicate with other neurons. In fact, the sensory neuron activated by the touch to the tail "talks" directly to a motor neuron which is responsible for the contraction of the tail. Motor neurons are nerve cells that control muscles. So when we touch the tail, the sensory neuron "talks" to the motor neuron and the motor neuron causes the muscle of the tail to contract.
When most people think about the concept of memory, they think of declarative memory. Declarative memory means that you can declare a fact that you know, or recall events, names and situations from the past. This type of memory is what you use when taking a test. However, memory, like test-taking, is not simple at all. Declarative memory is only one form of memory, called associative memory. There are also other forms of memory known as nonassociative memory.
Nonassociative memory includes two simple forms of learning called habituation and sensitization. Habituation is defined as a decrease in response to a repeated stimulus. For example, if you have ever noticed a smell when your nose first comes in contact with the odor, but some time later, you don't even notice that there is an odor present. In this case, your sense of smell has demonstrated habituation. You have stopped responding to the odor even though the odor is still present. This is a decreased responsiveness to the stimulus.
On the other hand, the other form of nonassociative learning, sensitization, is the opposite of habituation. Sensitization is an increased responsiveness For example, if you've ever been cut somewhere on your body and rubbed your hand across the cut, the skin around the cut is more sensitive than normal. This is because your skin has been sensitized. Normally, rubbing the skin would produce a mere sensation of touch, but this same touch to sensitized skin causes pain.
Aplysia are capable of demonstrating both sensitization and habituation. What does a sea slug have to remember? One of the most important things a sea slug must do is to learn how to survive in an ocean full of predators ready to eat them. Two main predators pursue sea slugs, crabs and lobsters. If you've ever been on the receiving end of a crab claw at the beach, you know just how painful this can be. Well, think how much more traumatic it is for a small, soft-bodied sea slug. Whether a sea slug becomes dinner or escapes to live another day will depend on how well the slug learns about and responds to its predators
A sea slug responds to its predators by reflexively withdrawing the portion of the body that is being pinched. This reflex is very similar to our knee-jerk reflex. You know when the doctor hits your knee with a hammer and you are unable to prevent your foot from kicking out? Reflexes are not under conscious control and occur very rapidly. It is possible for these reflexes to undergo sensitization or habituation. A crab claw can easily sensitize a slug's reflexes to future attacks. If the slug responds more rapidly to later attacks, it has a better chance of surviving. The basic component of any reflex is simply a sensory and motor neuron. Neurons are individual brain cells. A sensory neuron is responsible for sensing things in the environment, such as a touch, while a motor neuron is responsible for causing muscles to contract so that animals can move. When a crab claw touches or pinches a sea slug, the sensory neuron senses the claw and tells the motor neuron to contract the muscle so that the slug can escape. Sensitization of this simple reflex helps the slug to more quickly make its escape.
One of the most fascinating areas of Neuroscience research today uses Aplysia to study what changes occur at the level of individual sensory and motor neurons to bring about sensitization or learning. One of the theories that our laboratory is exploring is that primitive forms of learning may have evolved from basic reflex pathway modulation from painful stimuli. More simply, one of the first memories ever formed may have been a memory of a painful stimulus, such as an attack from a predator
http://www.mni.mcgill.ca/nm/1997f/en/aplysia.htm
You're at the movies watching a horror film. Goose bumps rise. The heartbeat quickens. Someone behind you abruptly lays a hand on your shoulder...
Why do we become hyper-sensitive at such moments and jump?
"We're talking about an incredibly complicated cognitive decision which we are nowhere near to understanding," remarks Dr. Wayne Sossin, a microbiologist who studies learning and memory at the Montreal Neurological Institute. A young Canadian who joined the MNI biology Unit two years ago, he has already been the principal author of 11 papers in some of the most prestigious international journals. His approach to "incredibly complicated" problems of cognition takes its cue from the method by which DNA was discovered almost half a century ago.
"Biologists had tried to understand life for hundreds of years and never made any progress," he mused recently in his seventh-floor laboratory. "Forty or fifty years ago, physicists came along and said, "Look, you're setting up too complicated a problem. You have to simplify it before you start."
Simplifying meant looking at organisms smaller than human beings. "It's from experiments with bacteria that we understood what DNA is," notes Sossin. Sossin experiments on creatures somewhat larger than bacteria, but which are tiny nonetheless: Aplysia californica, a slug-like marine mollusc, has become a favorite animal in behavior experiments.
"One of the reasons I chose Aplysia is that its neurons are quite large, says Sossin. "One can record from them much more easily than one can record from the mammalian neuron. Also, in the brain of a mollusc, there are particular neurons which are immediately identifiable, and which you can identity from animal to animal."
Scientists studying Aplysia have already learned a good deal about its neuronal behavior: for one thing, its neuronal system is plastic - it changes and adopts over time.
Much has been learned, too, about the exchanges that occur between Aplysia's nerve cell endings at the gap connecting them called the synapse. Sossin acknowledges that such background work has proved extremely useful.
"It helps to work in the same system in which a lot of the behavioral and synaptic changes have been made because one can then make correlations between the molecular level, the biochemical level and the cellular level - and eventually the behavioral level." All of which says that research on the slow-moving Aplysia has been very successful.
"It's probably the only system where an identifiable change in the connections between two identified neurons is known to have something to do with the behavior at the animal," says Sossin.
A good example comes from the neurons involved when Aplysia draws back as a result of some threatening stimulus.
"It's actually quite a complicated circuit that controls this, but when one just looks at the simplest part of the circuit - the sensory neuron for touch - it's connected directly to the motor neuron for withdrawal. Normally that connection is weak, and a simple touch does not cause the motor neuron to fire. When that connection gets made much stronger, then a touch causes the motor neuron to lire and the slug withdraws."
Scientists postulated that Aplysia's behavior changes because the connection between the sensory and motor neurons - the synaptic point at which an electrical impulse stimulates a chemical discharge - becomes stronger.
"In fact, it does get stronger," observes Sossin. "Now, there are many other neurons involved in a complex circuit, but that particular connection does contribute to changes in the animal's behavior. That's a powerful paradigm to work in because now we know that this neuron gets bigger in a behaviorally-relevant way."
Sossin aims to discover why this happens.
"It's a system you can look at on many different levels," he explains. "You can take those neurons and put them in a dish and they'll still make a connection between each other. And if you mimic the sensitizing stimulus with a drug such as serotonin, which Aplysia uses to signal a sensitizing episode, then you can see that the connection gets stronger in both the short term - a period of minutes - or in the long term - for days."
Such an observation has implications for Sossin's main focus of research short and long-term memory. But working with Aplysia has its shortcomings. It's not clear, for example, whether Aplysia has any long-term memories at all.
"Aplysia has a number at behaviors that are encoded genetically. They're not learned," remarks Sossin. "You don't teach a chicken how to lay an egg."
Another popular system for studying the molecules involved in learning and memory is the hippocampal slice, a thin section of the brain which is taken out of the hippocampus of a rat. The hippocampus is thought to he involved in learning and memory. Many of the neuronal changes which one observes in the hippocampus are similar to those in Aplysia, but for researchers such ac Sossin there are drawbacks.
"The hippocampal system is more complicated, and its link to behavior is not strong," he says. "It's not clear what it means when two neurons in the hippocampus are strengthened."
http://employees.csbsju.edu/hjakubowski/classes/ch331/signaltrans/olsignalkinases.html (my last section on Kinases)
Another Look at Neurotransmission and Ion Channels
You may have noticed above that some signaling molecules regulated by kinases (β-adrenergic and some olfactory signals by PKA and acetylcholine by PKC for example) are neurotransmitters. In our previous guide, we discussed how neurotransmitters can act as signals to open ion-specific, ligand-gated membrane channels, which change the transmembrane potential. In other words, the neurotransmitters gate the channels directly. Typical examples of channels directly gated by neurotransmitters are the acetylcholine receptor in neuromuscular junctions and the Glu, Gly, and GABA receptors in the central nervous system. These receptors are multimeric proteins. Receptors with direct gating of ion flow are fast, with activities that last milliseconds, and are used in eliciting behavioral responses.
However, ion channels can also be gated indirectly when the neurotransmitter binds to its receptor and leads to events which open an ion channel that is distinct from the receptor. In this case, the occupied receptor communicates to an ion channel indirectly through a G protein. Example of this indirect gating of ion channels include the serotonin, adrenergic, and dopamine receptors in the brain. These receptors are classic single protein serpentine receptors with 7 transmembrane helices, and intracellular domains that can interact with G proteins as described above to increase second messenger levels (cAMP, DAG) in the neuron. These can either activate kinases in the cell, which phosphorylate ion channels to either open or close them, or can bind directly to the channel and modulate its activity through an allosteric conformational change. In some cases the G protein directly acts on the ion channel. These different ways are described below. In contrast to direct gating, receptors that indirectly gate ion channels have activities that are slow and last seconds to minutes. These receptors are usually involved in modulating behavior by changing the excitability of neurons and the strength of neural connections, hence modulating learning and memory. These changes can occur in many ways:
Phosphorylating ion channels: Receptors that act through a second messenger system can change ion channel activity by activating kinases which phosphorylate the channels. This may:
open the channel normally closed at the resting potential and produce an effect like gating.
close a channel usually open at the resting potential (such as nongated K channels which when closed would depolarize the cell and make it more excitable).
Gα interaction with ion channels:
the Ga subunit of the G protein interacts with K channels after stimulation of the CNS Acetylcholine receptor, opening the channel and hyperpolarizing the cell
Second messenger interaction with ion channels:
cGMP opens cation channels in retinal cells after activation of the photoreceptor by photons
cAMP opens cation channels in olfactory cells after activation of the olfactory receptor by odorants.
Second messenger interaction with proteins other than ion channels (usually different receptors):
the β-adrenergic receptors are phosphorylated by PKA and PKC (activated by stimulation of a different neurotransmitter receptor linked through a G protein to produce increased levels of second messengers cAMP and diacylglycerol). When phosphorylated, the β-adrenergic receptor, itself activated through G protein) can't bind Gs. This in affect attenuates the response of the β-adrenergic receptor to its own neurotransmitter which leads to desensitization to that signal.
Second messenger regulates gene expression:
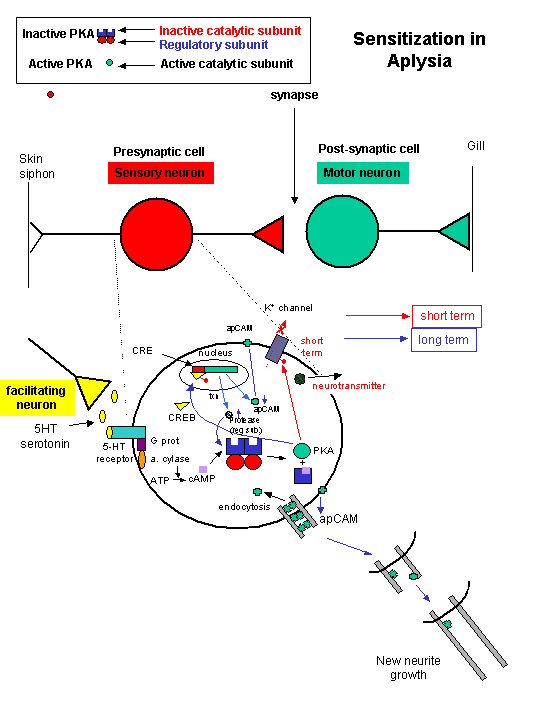
cAMP activated PKA can phosphorylate an inactive transcription factor in the cell, which then can bind to a section of DNA called the cAMP Response Element (CRE), which is upstream of certain genes, leading to the transcription of the genes. The transcription factor is called CREB for cAMP Response Element Binding protein. Example: tyrosine hydroxylase (a monooxygenase) is involved in the synthesis of epinephrine and norepinephrine. The activity of this protein is increased when it is phosphorylated by PKA. Hence its activity can be increased quickly by this modification of the already present protein. If an animal is subjected to severe or long term stress (cold or immobilization), presynaptic cells with norepinephrine will be stimlulated to release the neurotransmitter. This requires continual synthesis of the neurotransmitter by the presynaptic cell. The increase in synthesis of this neurotransmitter is caused by the presynaptic cell being stimulated by another neuron, whch leads to increased levels of cAMP, and ultimately activation of CREB which increases transcription of the hydroxylase gene.
http://hebb.mit.edu/courses/9.03/lecture3.html
9.03 Neural Basis of Learning and Memory: Lecture 3
Sensitization and Conditioning in Aplysia: Cellular mechansisms
Segue to the basic principles of second messenger processes
receptor ->G protein -> primary effector -> second messenger -> secondary effector
Examples of primary effectors are adenylyl cyclase producing cAMP, phospholipase C (PLC) producing IP3 and DAG, and phospholipase A (PLA) activating arachidonic acid. Either the G protein or the second messenger then act on a channel to modify ionic across the cell membrane.
Protein kinases
Back to sensitization
- seretonin binding to the 5-HT receptor activating a G protein
- G protein activates adenylyl cyclase
- Adenylyl cyclase increases the activity of cyclic AMP
- cyclic AMP activates a cAMP dependent protein kinase
Long-term facilitation acts through the same cAMP signalling system.
Second messengers can induce new protein synthesis by regulating gene expression.
Segue to gene regulation
Back to sensitization
Classical conditioning of gill withdrawal
Related mechansisms are involved in the classical conditioning of the withdrawal response.
The unconditioned stimulus (US) of tail shock produces the UCR of gill withdrawal.
Preceding the tail shock with stimulation of another sensory input (mantle shelf) facilitates the response to mantle stimulation.
The mechanism involves the increase of seretonin activited adenylyl cyclase activity (triggered by tail shock) in the presence of Calmodulin. Calmodulin is dependent on Ca++ which results from activation of Ca++ channels by sensory stimulation (mantle).
http://clem.mscd.edu/~bashamm/learning/aplysia.htm
Figure below shows neurons involved in habituation
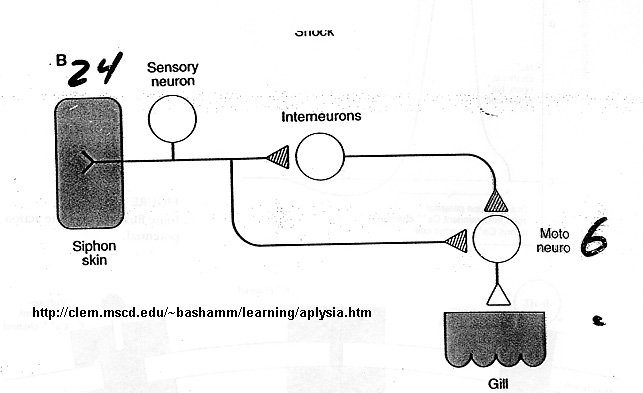
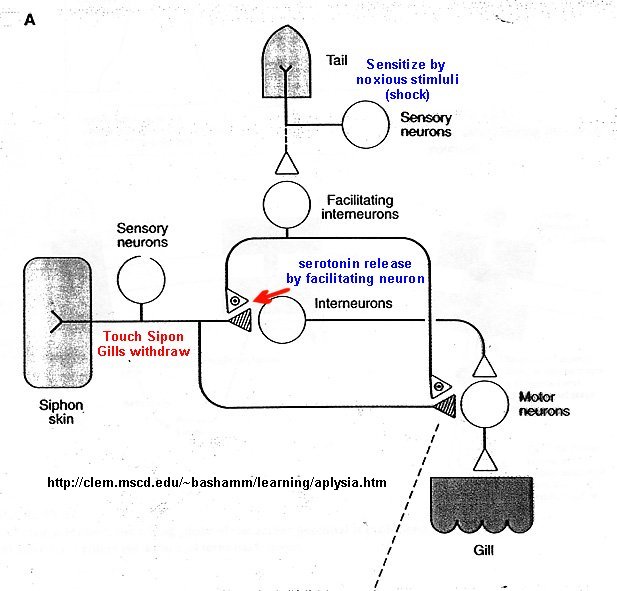
Figure below shows neurons involved in sensitiziation
http://www.hhmi.org/research/investigators/kandel.html
Summary: Eric Kandel's lab is studying explicit memory storage (the conscious recall of information about people, places, and objects) in mice and implicit memory storage (the unconscious recall of perceptual and motor skills ) in the snail Aplysia. In Aplysia, the lab has focused on the implicit memory for sensitization, a simple form of learned fear, and the mechanisms for achieving synapse-specific anatomical changes. In mice, they have examined the synaptic mechanisms contributing to memory for space, a complex form of explicit memory storage.
Cognitive psychological studies in patients with brain lesions and normal subjects indicate that there are two distinct types of memory storage: the conscious recall of information about people, places, and things (explicit memory), and the unconscious use of information about motor skills and perceptual strategies (implicit memory). The existence of two very different forms of memory storage raises the question: To what degree do these different memory processes share molecular steps? One clue to shared mechanisms comes from the study of stages in memory storage. Both implicit and explicit forms of memory have at least two phases of storage: a short-term phase lasting minutes and a long-term phase lasting days or longer. These two phases differ not only in their time course but also in their molecular mechanisms: long-term memory but not short-term memory requires gene expression and the synthesis of new protein. Detailed studies of implicit memory in Aplysia and Drosophila and preliminary studies of explicit memory in mice suggest that the requirement for protein synthesis that characterizes long-term memory is reflected on the cellular level by changes in the strength of synaptic connections. These changes result from the activation of a cascade of gene expression (beginning with the activation of CREB1), which leads to the growth of new synaptic connections.
Earlier molecular studies of gene expression related to memory storage focused on the identification of positive regulators. Recent studies in Aplysia have revealed a surprising finding: long-lasting forms of synaptic plasticity related to simple implicit storage require not only the activation of positive regulatory mechanisms that favor memory storage but also the removal of inhibitory constraints that oppose memory storage. By analogy to tumor-suppressor genes, which restrain cell proliferation, we have called these inhibitory constraints on memory storage, which restrain synapse growth, memory-suppressor genes. We now have found that there also are memory-suppressor genes for complex explicit memory in mice.
Memory-Suppressor Genes for Explicit Memory Storage in the Mammalian Brain
The study of explicit memory has focused on positive regulators such as kinases.
By contrast, relatively little attention has been paid to phosphatases.
Phosphatases are likely to have signaling roles in synaptic plasticity and
memory as important as those of kinases, if only because of their antagonistic
relationship: the ability of phosphatases to inhibit the action of kinases.
Furthermore, most cellular models of learning postulate erasure mechanisms
designed to counteract long-lasting synaptic enhancement. To examine the role of
specific phosphatases in synaptic plasticity, we focused our initial efforts on
calcineurin (protein phosphatase 2B, PP2B). This enzyme is thought to be the
first step in a phosphatase cascade initiated by Ca2+
influx through the NMDA receptor, the key receptor activated in learning-related
synaptic plasticity.
To examine further the role of phosphatases in synaptic plasticity and in memory storage, we have carried out two types of experiments: (1) we overexpressed in the mouse forebrain a truncated form of calcineurin Aα, and (2) we expressed a dominant negative inhibitor of calcineurin. To assure that a genetic overexpression of calcineurin is both spatially restricted and temporally regulated, we used the promoter of the calcium-calmodulin�independent kinase, which restricts expression to the forebrain, and tTA and rtTA, the tetracycline system, to obtain temporal control of gene expression and to show reversibility of the phenotype. Using the rtTA system we found that doxycycline induced maximal gene expression in neurons of the hippocampus within six days and that this expression can be reversed by removal of doxycycline. This doxycycline-induced expression of calcineurin reversibly impaired an intermediate (protein kinase A [PKA]-dependent) form of long-term potentiation (LTP) in the hippocampus as well as the transition from short- to long-term storage of spatial memory. This reversibility shows (1) that the transgene did not produce these effects on LTP and on memory storage by affecting development and (2) that calcineurin can act as an inhibitory constraint on PKA-dependent processes and can down-regulate pathways supporting synaptic plasticity and memory.
To explore this finding further, we next carried out the reverse experiment, the reversible expression of an inhibitor of calcineurin. The transient reduction of calcineurin activity resulted in (PKA-dependent) facilitation of LTP both in vitro and in vivo. This facilitation persists over several days in the awake animal, and is accompanied by enhanced learning and strengthening of short- and long-term memory on several hippocampal-dependent spatial and nonspatial tasks. Again the improvements in both LTP and memory were reversed fully upon suppression of the transgene. Together, the results of these two experiments demonstrate that endogenous calcineurin is an inhibitory constraint on synaptic plasticity and memory storage.
Synapse Specificity: A Function for Local Protein Synthesis in Memory
Storage
The requirement for transcription during long-lasting forms of synaptic
plasticity related to both explicit memory in mice and implicit memory in Aplysia
has raised the question of whether the critical unit of synaptic plasticity is
the nucleus or the synapse. If it is the synapse, then there must be a set of
mechanisms that allows the products of gene expression to alter synaptic
strength at some synapses but not at others on the same cell. Each of the
approximately 1011 neurons in the brain makes on average 1,000
synaptic connections on its population of target cells. It has been thought
that, to maximize information processing, the critical cellular unit of synaptic
plasticity is the individual synapse. Although attractive, this idea has never
been tested directly at the level of individual neurons, because there have not
been appropriate cellular systems for exploring long-term changes in the
synaptic strength of different terminals of an individual neuron. To address
this question, Kelsey Martin, Andrea Casadio, and I have developed a new culture
system consisting of a single Aplysia sensory neuron with a bifurcated
axon making synaptic contact with two spatially separated target motor neurons.
By perfusing serotonin onto only one of the synaptic connections we found that a
single axonal branch is able to undergo both short-term and long-term
synapse-specific facilitation. The long-term synapse-specific facilitation
produced by repeated local exposure to serotonin is dependent on CREB-mediated
transcription and involves the growth of new synaptic connections at the
serotonin-treated, but not at the untreated, branch. This culture system also
demonstrates synaptic "capture"�the ability to establish the
long-term process at a second branch receiving a subthreshold stimulus once
transcription-dependent facilitation has been established at another synapse in
the same cell. Thus, when a single pulse of serotonin (which produces only
transient, short-term facilitation) is applied to one synapse immediately
following five pulses of serotonin to the other synapse, long-lasting
facilitation and the growth of new connections occurs at both branches. The
establishment of synapse-specific long-term facilitation requires rapamycin-sensitive
local protein synthesis in the presynaptic, but not the postsynaptic, cell. To
study this local protein synthesis, we have begun to use sensory cell processes
deprived of their cell bodies, and have found that basal protein synthesis is
stimulated threefold by exposure to serotonin.
Summary: Changes in Sensory Cells in Habituation
http://www.fmrib.ox.ac.uk/~stuart/thesis/chapter_3/section3_2.html B
Brain Sensory Areas
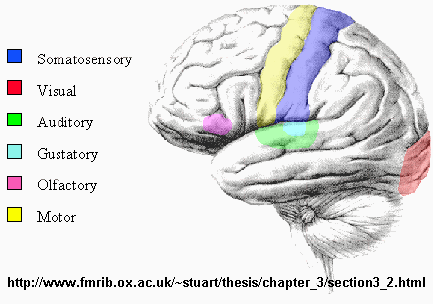
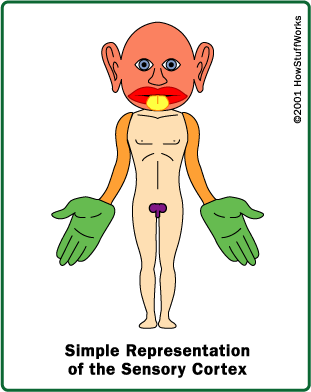
Somatosensory Map - Homunculus
Increased auditory cortical representation in musicians
Recent Articles:
Dudai, Y. Neruobiology: Fear Thou Not. Nature. 421, pg 325 (2003). Neurochemistry of fear forgetting and PTSD.
Gernoux et al. Protein Phosphatase 1 is a molecular constraint on learning and memory. Nature. 418, pg 970 (2002)
Silva and Josselyn. The Molecules of Forgetfulness. Natuer. 418, pg 929 (2002)
{kind=link}