{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
![]() Chime Molecule Modeling: Acetylcholine Receptor Pore
|
Jmol
(1OED)
Chime Molecule Modeling: Acetylcholine Receptor Pore
|
Jmol
(1OED)
02/17/2008
Neurobiochemistry is one of the most explosive areas of biological research. Scientists are now starting to unravel the molecular bases for memory, cognition, emotion, and behavior. The next decades will bring truly revolutionary understanding of brain chemistry and along with it the potential to alter human mood, memory, and to treat mental illness such as schizophrenia much more effectively. The human brain, with about 100 billion neurons (each which can form connections - synapses - with 1000 to 10,000 other neurons ) and associated glial cells (10-50 times the number of neurons) can be considered one of the most complex structures in the universe. This section will explore the biology and chemistry of neurons.
THE RESTING POTENTIAL AND ACTION POTENTIALS
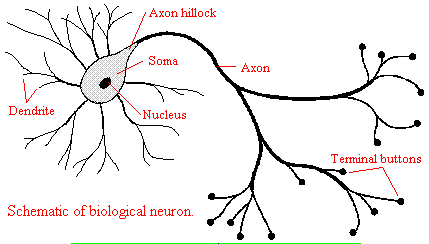
Neurons consist of a single, nucleated cell body with multiple signal-receiving outgrowths (dendrites) and multiple-signal sending outgrowths (axons) which end in a terminal button. These interact through the synapse with dendrites on other neurons.
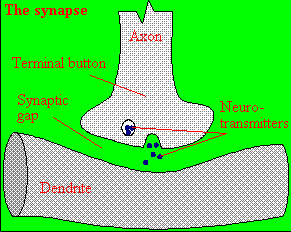
A presynaptic neuron can stimulate an adjacent postsynaptic neuron by releasing a neurotransmitter into the synapse between the cells, which binds to a receptor in the membrane of the post-synapatic cell, stimulating the cell. We will discuss the events which cause the post-synaptic cell to "fire", but we will not discuss the immediate events which lead to the release of neurotransmitter by the presynaptic neuron.
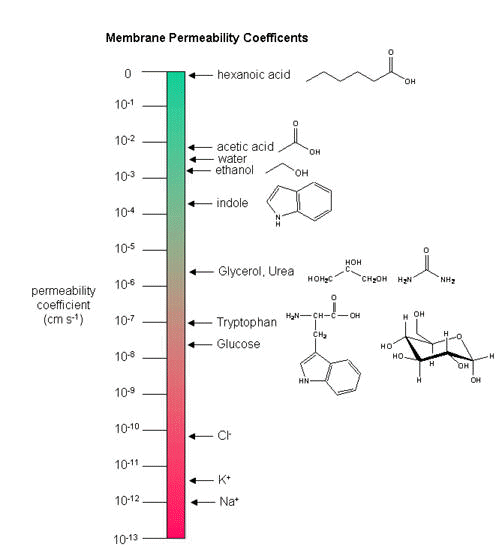
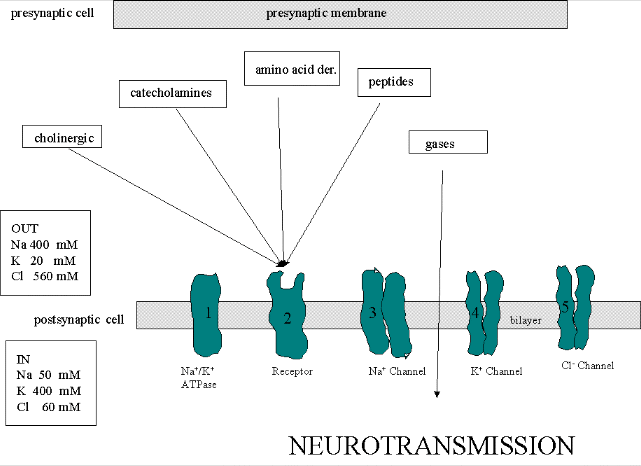
Neurons (as do all cells) have a transmembrane voltage difference or potential across the membrane. Transient changes in the membrane potential are associated with neuron activation or inhibition. This arises in part due to the imbalance of sodium and potassium ions across the membrane which were established by a protein , Na+-K+-ATPase, in the membrane. This protein transfers 3 Na+ ions out of the cytoplasm for every 2 K+ ions it transports in, which generates a transmembrane potential. Likewise Cl- has a much higher level outside the cell. Membrane potentials are determined not only by the size of the ion gradients across the membrane, but also the differential permeability of membranes to ions. Synthetic bilayer membranes are not very permeable to ions. This should follow from your understanding of intermolecular forces: ions are not stabilized by nonpolar molecules and are not soluble in nonpolar solvents. the table below shows permeability of various ions to a liposome which is a bilayer without membrane proteins.
Ion permeability of phosphatidyl serine vesicles
| ION | PERMEABILITY (cm/s) |
| sodium | <1.6 x 10-13 (lowest) |
| potassium | <9 x 10-13 |
| chloride | <1.5 x 10-11 (highest) |
Now check out the table below which shows the concentrations of ions inside cells and outside (for example in the blood) and their permeabilities to mammalian bilalyer membranes.
Typical ion concentrations and permeabilities for mammalian membranes.
| Ion | Cell (mM) | Blood (mM) | Permeability (cm/s) |
| potassium | 140 | 5 | 5 x 10-7 |
| sodium | 5-15 | 145 | 5 x10-9 |
| chloride | 4 | 110 | 1 x 10-8 |
| X- (neg. macromol.) | 138 | 9 | 0 |
How can we account for the markedly greater permeabilities of ions (1000x to 1,000,000 x) in mammalian cell membranes compared to synthetic lipid vesicles? Glucose also has a greater permeability through red blood cell membranes than through synthetic liposomes because of a membrane receptor that allows facilitated diffusion across the membrane and down a concentration gradient.
The same thing is true of ion permeabilites in intact biologicial membranes. These membranes have several types of selective ion channels (nongated - always open, and gated - open only after specific shape changes in the protein). The nongated channels dramatically increase the permeability of membranes to ions, as the glucose transport protein increased the permeabilty to glucose. Ion channels in nerve and muscle can move ions across the membrane at a rate up to 109/s..
The Transmembrane Potential
How is the transmembrane potential formed? Both glial cells (which function as protectors, scavengers, and
feeder for brain neurons) and neurons have transmembrane potentials. First
consider glial cells.
Glial Cells
The transmembrane ion gradients for ions are established, in part, through the action of ion-specific ATPases, such as we discussed with the Na/K ATPase. This transporter ejects 3 sodium ions from the inside of the cell for every 2 potassium ions in transports in, all against a concentration gradient. This makes the inside more "negative" than the outside, which contributes to the transmembrane potential. But note also that there is a Cl ion gradient across the membrane also. If another transporter moves Cl- ions to the outside of the cell in equal amounts as for Na+ ions, no charge imbalance would exists across the membrane and no transmembrane potential would exist. In addition, might not any initial charge imbalance across the membrane, which would lead to a transmembrane potential, collapse as the ion gradient collapses as sodium flow back across the membrane down its concentration gradient and potassium ions flow out?
Two things must occur for a membrane potential to be formed and be stable.
With respect to ion channels, glial cells appear to have only a non-gated potassium channel, which allows the outward flow of potassium ions down the concentration gradient. The inside will then have a net negative charge since impermeable anions remain. Sodium can't get from the outside to the inside through a channel. The concentration gradient causes this outward flow of potassium ions. As more ions leave, the inside gets more negative, and a transmembrane potential which resists further efflux of potassium develops. Eventually they balance, and the net efflux of potassium stops. The resting transmembrane potential reaches -75 mV .
Neurons
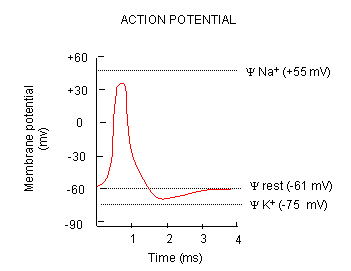
We can easily measure the actual transmembrane potential of cells. Varying the outside sodium and potassium concentrations would change the experimental transmembrane potential,. The experimental resting potentials of glial cells always matched the theoretical potassium potentials, supporting the view that the transmembrane potential was associated only with open, nongated potasisum channels. This was not observed with neurons, suggesting that channels other than for potassium were open. It became clear that nerve cells were permeable not only to potassium, but also to sodium and chloride. How do these work in establishing the resting potential? Consider the simplest case when just potassium channels are present, along with an unequal distribution of other ions. Now add some sodium channels. Two forces act to drive sodium into the cell - the concentration gradient, since sodium is higher on the outside, and the membrane potential since the inside of the cell is negative. The equilibrium potential of a cell if it were only permeable to sodium is +55 mv, so there is a great electrical drive for sodium to enter through the nongated, open sodium pores we just added. As sodium enters, the cell starts to "depolarize" and have a more positive voltage. However, since in our example, there are many more open potassium channels, the resting potential deviates only a small amount from the potassium potential, since as the potential becomes more positive, more potassium flows out down the concentration gradient. Eventually the enhanced potassium efflux equals the sodium influx, and a new resting membrane potential of -60 mV is established, which is typical of neurons. In the resting cells, the passive fluxes of sodium and potassium ions are exactly balanced by the active fluxes of these ions mediated by the Na/K ATPase.
BINDING OF NEUROTRASMITTER TO RECEPTORS: CELL ACTIVATION
What happens when a neurotransmitter binds to a receptor on the post-synaptic cell?
In the central nervous system, life is pretty complicated:
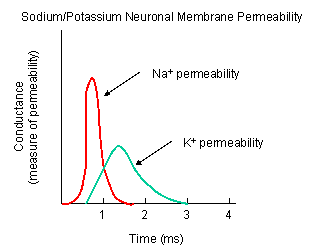
Now what happens when a neurotransmitter binds to the receptor on the post-synaptic cell? A depolarization occurs (mediated by conformational changes in the transmitter-receptor complex) raising the membrane potential from the initial steady level. What happens next depends on the identity of the post synaptic cell. In a neuron, the rising potential triggers an action potential by opening voltage-gated sodium channels. The potential rises to about + 35 mV, but does not reach the Na ion equilibrium potential, because the high positive potential opens a voltage-gated potassium channel. The potential then falls until it reaches the K ion equilibrium potential where the cells is hyperpolarized. It slowly then relaxes back to the resting potential of -60 mV. This wave of changes in potential sweeps down the post-synaptic cell membrane and is the basis for the "firing" of the neuron.
The following incredible animations come from: Neurobiology by Gary Matthews.
Animations from Biology by Raven et al.
THE PROTEIN PLAYERS
To understand how neurons we need to consider five membrane proteins:
Chime Molecule Modeling: Acetylcholine Receptor Pore | Jmol (1OED)
![]() Chime Molecule Modeling: Potassium
Channel |
Jmol
Chime Molecule Modeling: Potassium
Channel |
Jmol
Excitatory Neurotransmitters in the Brain:
Glutamate is a major excitatory neurotransmitter in the brain. Four types of glutamate receptors are found in the central nervous system. They differ in the nature of neurotransmitter which bind to the receptor and which act as agonists. Excessive amounts of glutamate are neurotoxic.
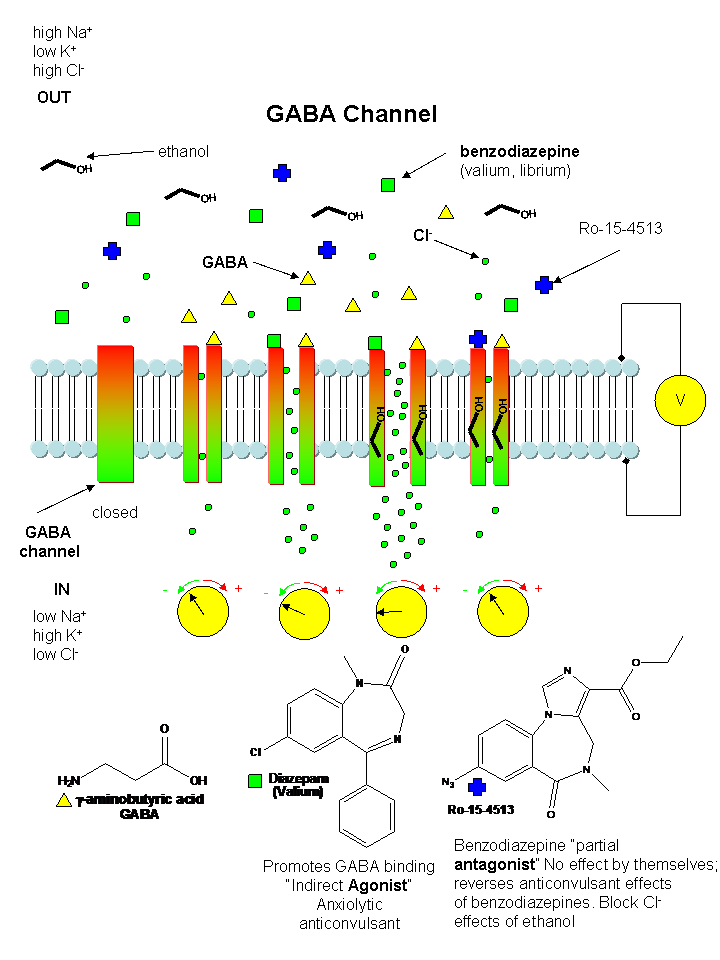
Inhibitory Neurotransmitters:
The main inhibitory neurotransmitters are GABA (gamma-aminobutyric acid )and glycine. They bind to neurotransmitter-gated chloride channels, which when open hyperpolarize the membrane (make the transmembrane potential more negative) and hinder neuron firing. Benzodiazepines (like Valium and Librium - anti-anxiety and muscle-relaxing agents) and barbituates (like phenobarbital-hypnotics) bind at sites other than site where GABA binds and potentiate (increase) the binding of each other and GABA.
![]() Chime
Molecule Modeling: Chloride
Channel |
Jmol
Chime
Molecule Modeling: Chloride
Channel |
Jmol
Summary:
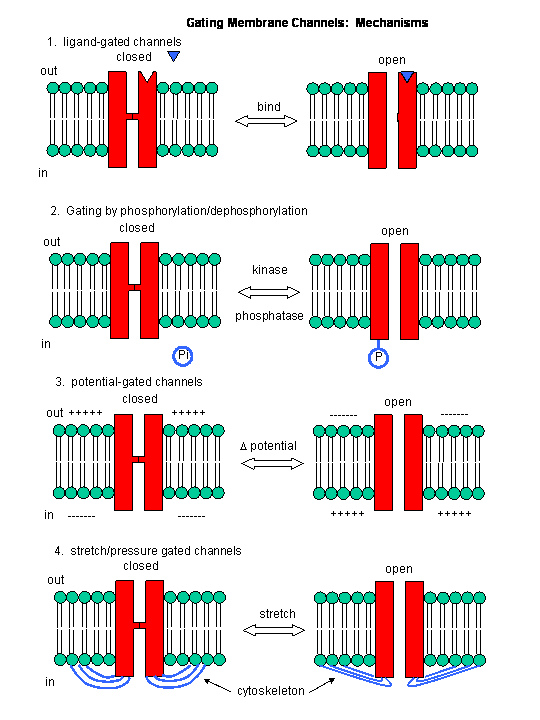
In summary, neurotransmitter and voltage gated channels allow changes in the polarization of the membrane. Other mechanisms can also lead to changes. Membrane proteins can be phosphorylated (using ATP) by protein kinases in the cell, leading to a change in the conformation of the membrane protein, and either an opening or closing of the channel. Channels linked to the cytoskeleton of the cells can also be opened or closed through stretching. Other stimuli that gate channels are light (through photoisomerization-induced conformational changes), heat, and cold.
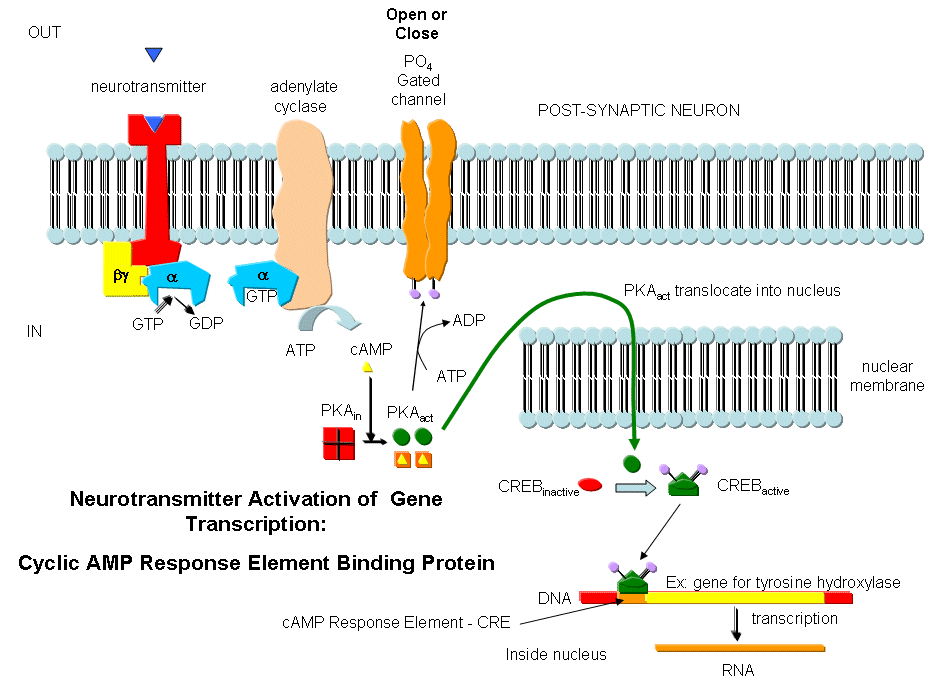
Neurotransmitters can act as signals to open ion-specific, ligand-gated membrane channels, which change the transmembrane potential. In other words, the neurotransmitters gate the channels directly. Typical examples of channels directly gated by neurotransmitters are the acetylcholine receptor in neuromuscular junctions and the Glu, Gly, and GABA receptors in the central nervous system. Receptors with direct gating of ion flow are fast, with activities that last milliseconds, and are used in eliciting behavioral responses.
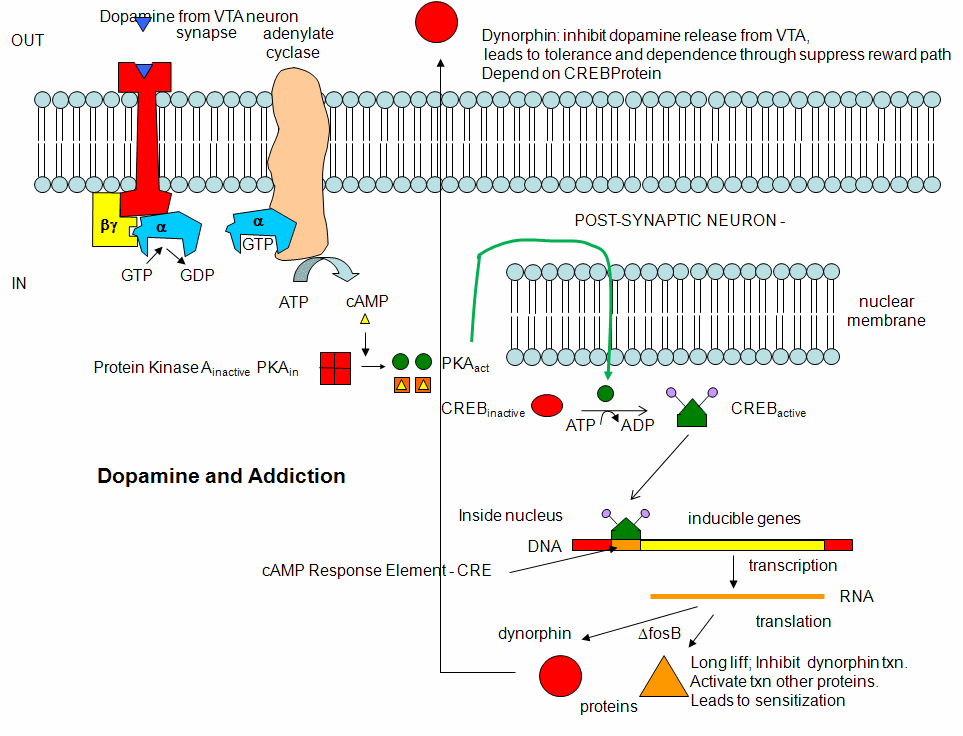
However, ion channels can also be gated indirectly when the neurotransmitter binds to its receptor and leads to events which open an ion channel that is distinct from the receptor. In this case, the occupied receptor communicates to an ion channel indirectly through activation of kinases which can phosphorylate other protein including other neurotransmitter receptors. Example of this indirect gating of ion channels include the serotonin, adrenergic, and dopamine receptors in the brain. These receptors, when they bind neurotransmitters, lead to the increase of second messenger levels (such as cAMP) in the neuron. This can either activate kinases in the cell, which phosphorylate ion channels to either open or close them, or can bind directly to the channel and modulate its activity through a direct shape change. In contrast to direct gating, receptors that indirectly gate ion channels have activities that are slow and last seconds to minutes. These receptors are usually involved in modulating behavior by changing the excitability of neurons and the strength of neural connections, hence modulating learning and memory.
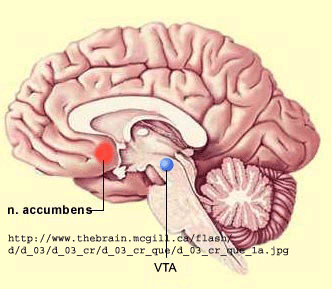
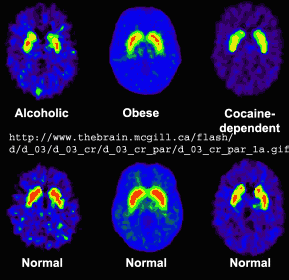
Alcohol and Drug Effects on Neurons
You should now have enough background to read some recent scientific articles on the action of alcohol and other drugs on neurons. Please refer to the class schedule and WebCT for specific reading assignments. I will add additional summarizes below from recent studies if warranted.
Alterations of potassium channel have recently been implicated in the inhibitory effects of ethanol. Davies, McIntire, et al. studied effects of ethanol on the round worm C. elegans, believing that alcohol-mediated inhibition of neural activity would be conserved across species. Previously it has been shown that the dose required for behavioral changes is similar for both invertebrates and vertebrates. Ethanol seems to affect many different proteins that would lead to synaptic inhibition, including GABA and glutamate channels and potassium channels. Many different gene products (dopamine D4 receptors, protein kinase C) are associated with increased sensitivity while others (nitric oxide synthase, dopamine D2) are associated with ethanol resistance. These changes suggests that complex pathways are involved in ethanol effects but they didn't isolate a specific target for its effects.
When C. elegans were exposed to ethanol for brief time periods, ethanol levels rose to values similar to levels seen in intoxicated drivers (0.1%). They isolated mutants that were resistant to the inhibitory effect of ethanol (which in this organism were observed as changes in movement and egg-laying behavior). These mutants affected a single gene, slo-1, homologous to the the slowpoke gene in drosophila. The gene in both organisms encoded a potassium channel, whose normal function is to repolarize neural membranes to their resting potential. The normal channel is activated by ethanol, which enhances K+ efflux, making the transmembrane potential more negative, which makes inhibits neural firing (the same outcome as when ethanol enhances Cl- influx through GABA channels). Mutants strains (resistant to alcohol), when transformed with slo-1+ regained ethanol sensitivity. Ethanol appears to directly activate the channel. This would lead to efflux of potassium ions from the worm, hyperpolarizing the neural cells, leading to inhibition of neural activity. Effects in C. elegans were observed at physiologically relevant ethanol, ranging from those that produce euphoria to sedation in humans.
Davies, A. et al. A Central Role of the BK Potassium Channel in Behavioral Response to Ethanol in C. Elegans. Cell, 115, pg 655 (2003)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}