Biochemistry Online: An Approach Based on Chemical Logic

CHAPTER 2 - PROTEIN STRUCTURE
D: PROTEIN FOLDING AND STABILITY
BIOCHEMISTRY - DR. JAKUBOWSKI
Last Update: 3/1/16
|
Learning Goals/Objectives for Chapter 2D: After class and this reading, students will be able to
|
D6. Protein Folding In Vivo
There are many differences between how a protein might fold or unfold in a cell compared to a test tube.
- The total concentration of all the proteins and nucleic acids in cells are estimated to be about 350 g/L, or 350 mg/ml. Most measurement in the lab are conducted in the range of 0.1 to 10 mg/ml
- Proteins are synthesized in cells from an N to C terminal direction. Hence the nascent protein, as it emerges from its site of synthesis (the ribosome), might fold into intermediate structures since not all of the protein sequence is yet available to direct folding.
- Proteins are synthesized in the cytoplasm, but they have to find their final place in the cell. Some end up in membranes, some must translocate across one or even two different membranes to end up in specific organelles like the Golgi, mitochondria, chloroplasts (in plant cells), nuclei, lysosomes, peroxisomes, etc. Do they translocate in their native state?
Additional evidence suggests that protein folding/translocation requires assistance (i.e. catalysis) in the cell.
- Mutant cells defective in certain proteins can lead to the accumulation in the cells of misfolded and aggregated proteins.
- eukaryotic genes (taken from higher cells which contain nuclei and internal organelles), when transferred into prokaryotes (bacteria, like E. Coli), can be expressed to form protein, but they often misfold and aggregate in the bacterial cells and form structures called inclusion bodies.
Hence recombinant proteins expressed in vivo have the same problems in folding as larger proteins in vitro. In both cases, conditions favor accumulation of nonnative proteins with exposed hydrophobic groups leading to aggregation. Aggregation also occurs in vivo when a protein is over-expressed or expressed at a higher temperature than normal. Why? Mutant cells have been selected that actually suppress inclusion bodies in vivo. This effect was mediated by a class of proteins which are expressed by the bacteria and other cells when their temperature is raised. The function of these proteins, called heat shock proteins (Hsp), was unknown until it was realized that they facilitate correct protein folding, in part, by binding to denatured proteins in the cells before they aggregate into inclusion bodies. Further studies discovered a large number of proteins that seem to facilitate protein folding and prevent aggregation in vivo. These proteins are now called molecularchaperones. They are classified on the basis of their molecular weight) and can be divided into at least two families, the Hsp-70 chaperone family and the chaperonin and Hsp 90 families as illustrated and summarized in the figure and text below.
Hsp-70 Family: This family includes DnaK/DnaJ and GrpE proteins in prokaryotes and immunolglobulin heavy chain binding protein (BiP) and alpha crystalline in eukaryotes. Alpha crystalline comprises 30% of the lens proteins in the eye, where it functions, in part, to prevent nonspecific, irreversible aggregates. These proteins have molecular weights of about 70K and :
- bind to growing polypeptide chains as they are synthesized on ribosomes.
- express activity as monomers.
- have ATPase activity - i.e. they cleave the phosphoanhydride ATP (which can drive reactions).
- bind short, extended peptides, which stimulates the ATPase activity
- release bound peptides after ATP cleavage
A figure showing
In prokaryotes, a protein called trigger factor (TF) binds in a co-translational process to proteins as they begin to emerge from the ribosome and catalyzes correct folding of about 70% of bacterial protein. The rest requires additional chaperones, including DnaJ and DnaK which bind proteins during synthesis in a cotranslational process. Upon interaction with the DnaJ-bound protein, DnaK hydrolyzes bound ATP, resulting in the formation of a stable complex between DnaJ and DnaK. GrpE, a nucleotide exchange factor for DnaK, facilitates the releases ADP from DnaK. Rebinding of ATP to DnaK then triggers the release of the substrate protein. This cycle repeats itself until the protein is fully folded. For about 20% of proteins in E. Coli, the DnaK/DnaJ/GrpE cycle leads to complete post-translational folding of proteins . Eukaryotes utilize an analogous set of proteins Hsp70 complex proteins including If folding is still incomplete after several rounds, the fully synthesized yet incompletely folded protein interacts with an amazing catalyst of protein folding, the chaperonin system.
Chaperonins- including chaperonin 60 (or GroEL in E. Coli) and chaperonin 10 (or GroES in E. Coli) in chloroplasts, mitochondria and bacteria, and TCP-1 in eukaryotic cytoplasm.
These proteins:
- bind to proteins after they have left the ribosome or have been transported into organelles like mitochondria.
- express activity as multimers. GroEL consist of two stacks of rings of monomers, with 7 monomers in each ring (each monomer around 60K MW), forming a hollow cylinder. GroES consist of one single ring of 7 monomers (each 10K MW). The GroES complex forms a lid over one open end of the GroEL cylinder. Proteins can fold within the cavity in GroEL (lined with hydrophobic patches) without "fear" of aggregation. GroEL also binds and cleaves ATP, leading to conformational changes inside the barrel and hiding of the hydrophobic patches in Gro EL, which leads to the releases of the unfolded peptide. The process proceeds until the folding protein passes through the barrel and is released in its correct folded state. Do you find this amazing?
- bind nonnative proteins at the GroEL opening of a complex of GroEL and GroES, which has a large hydrophobic cavity.
- Molecular Chaperonins in Disease
GroEL has also been shown to bind in its hydrophobic cavity a fluorescent CdS semiconductor nanoparticle which can be released on addition and cleavage of ATP. There are two classes of chaperonins:
- Class I: Those found in bacteria, chloroplasts and mitochondria. The have structures analogous to GroEL (two rings of 7 identical monomers) and Gro ES.
- Class II: Those found in archebacteria and in the cytoplasm of eukaryotic cells. These contain two rings of 8-9 subunits which may not be identical.
![]() Jmol:
Updated GroEL/ES
Jmol14 (Java) |
JSMol (HTML5)
Jmol:
Updated GroEL/ES
Jmol14 (Java) |
JSMol (HTML5)
Other chaperons have proven to be of clinical significance. Hsp 90 is a chaperone that is expressed both in normal and tumor cells. It appears to have special importance in tumor cells in helping key proteins involved in malignancy (signal transduction proteins such as HER-2/ErbB2, Akt, Raf-1, Bcr-Abl, and p53) to maintain their shapes under conditions of drug exposure and the inherent genetic instability present in the cells. Drugs that bind to and inhibit Hsp90 appear to have much greater effect on tumor cells, making this protein a new chemotherapeutic target to treat cancer. Recent studies by Kamal et al. have shown the drug 17-AAG binds Hsp90 about 100 times as strongly in tumor cells than in normal cells. Hsp 90 appears to be complexed to other "co-chaperones" in the tumor cells which lead to higher drug binding affinity. The chaperone complex may actually induce the drug to adopt a different conformation. A comparison of chaperone catalyzed folding in prokaryotes and eukaroytes is shown below.
Figure: Comparison of cytosolic prokaryotic and eukaryotic chaperone pathways
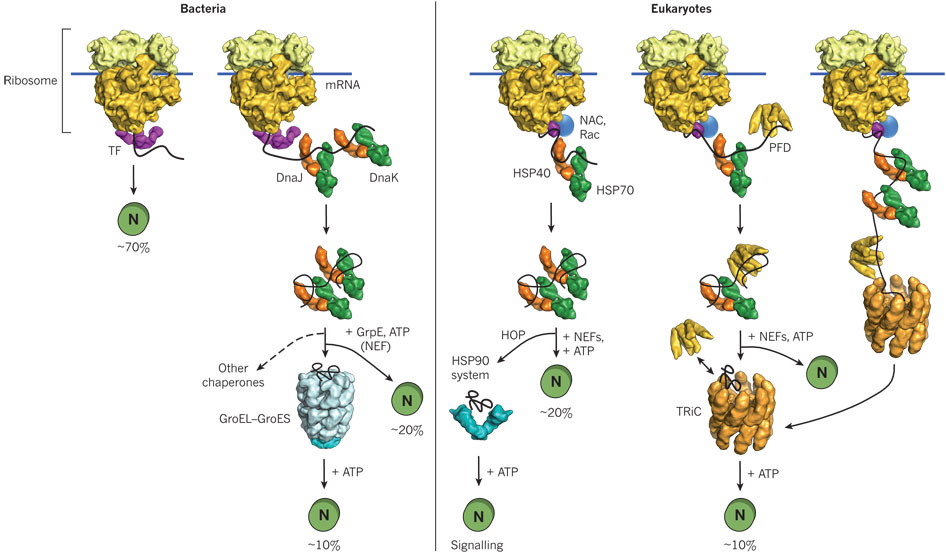
- Chaperone-assisted protein folding: Arthur Horwich (Yale/HHMI) Part 1A:
Additional Proteins Which Catalyze Protein Folding: Chaperons function to minimize protein aggregation, which increases the efficiency of the entire process. Other proteins in the cell actually catalyze specific steps. Here are two examples:
- Protein Disulfide Isomerase (PDI) - catalyzes the conversion of incorrect to correct disulfides. The active site consists of 2 sets of the the following sequence - Cys-Gly-His-Cys, in which the pKa of the Cys are much lower (7.3) than normal (8.5). How would this facilitate disulfide isomerization?
- Peptidyl Prolyl-Isomerase (PPI) - catalyses X-Pro isomerization, by a mechanism which probably involves bending the X-Pro peptide bond. How would this facilitate the process?
Many proteins have been found to possess PPI activity. One class is the immunophilins. These are small proteins found in the cytoplasm that bind anti-rejections drugs used to prevent tissue rejection after transplanation. The immunophilin FK506 binding protein (FKBP) binds FK506 while the protein cyclophilin binds that anti-rejection drug cyclosporin. The complex of cyclophilin:cyclosporin or FKBP:FK506 binds to an inhibits calcineurin, an important protein (with phosphatase activity) in immune cells (T cells) required for T cell function. In this case, immunophilin:drug binding to calcineurin inhibits the activity of the T cell, preventing immune attack on the transplanted tissue, preventing rejection. The immunosuppressant drugs (FK506 and cyclosporin) inhibit the PPI activity of their respective immunnophilin. The extent to which the PPI activity of cyclophiin is required for its activity is unclear, but it seems to be important for some of its biological effects.
![]() Jmol:
Updated Cyclophilin
Jmol14 (Java) |
JSMol (HTML5)
Jmol:
Updated Cyclophilin
Jmol14 (Java) |
JSMol (HTML5)
- Animation: Folding and Degradation of Proteins in vivo
As the site responsible for folding of membrane proteins and proteins destined for secretion, as well as the major site for lipid synthesis, the endoplasmic reticulum (ER) must be able to maintain homeostatic conditions to ensure proper protein formation. Plasma cells that synthesize antibodies for secretion as part of the immune activation, show large increases in protein chaperones and ER membrane size
The main pathway controlling ER biology is the unfolded protein response (UPR) signaling pathway. If demand for protein synthesis in the ER exceeds capacity, unfolded proteins accumulate. This ER stress conditions activates a protein called IRE1, a transmembrane Ser/Thr protein kinases (which phosphorylates proteins). IRE1 activates a transcription factor that controls transcription of many genes associated with protein folding in the ER. Another protein, ERAD (ER-associated degradation) which moves unfolded proteins back into the cytoplasm where they are degraded by the proteasome. Proteins involved in lipid synthesis are also activated as lipids are needed for membranes as the ER increases in size. If the stress can not be mitigated the signaling pathway leads to programmed cell death (apoptosis).
Schuck at al investigated the specific role and importance of UPR in the homeostasis of ER as modeled by the yeast Saccharomyces cervisiae. The UPR signaling pathway was analyzed using light and electron microscopy to visualize and quantify ER growth under various stress conditions. Western blotting procedures were performed to determine chaperone protein concentrations after stress induction and association with ER expansion after the ER was exposed to various treatment conditions. The authors found ER membrane expansion occurred through lipid synthesis since stress induction increased concentrations of proteins responsible for promoting lipid synthesis and expansion failed when the proteins were absent and lipid concentration was low. In addition, these lipid synthesis proteins were activated by the UPR signaling pathway. By separating ER size control and UPR signaling, they found that expansion occurred regardless of chaperone protein concentrations. However, if lipid synthesis genes were not available, raising the ER chaperone level helped alleviate stress levels in ER.
 Navigation
Navigation
Return to Chapter 2D: Protein Folding and Stability
Return to Biochemistry Online Table of Contents
Archived version of full Chapter 2D: Protein Folding and Stability

Biochemistry Online by Henry Jakubowski is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.