BIOCHEMISTRY - DR. JAKUBOWSKI
This material has not be updated for a few years.
INTRODUCTION
What better way to end the semester with a study of one of the hottest areas in signal transduction - programmed cell death or apoptosis. We have spent time discussing how cells receive external signals from the surrounding environment, which cause a cellular response such as differentiation, proliferation, secretion, etc. We have also seen how important it is for cells to be able to adapt to changes and how exquisitely controlled the process of signal transduction is. One appropriate response to a signal is for the cell to commit suicide - presumably for the good of the organism. Over the past decade it has become very clear that programmed cell death is encoded in the genome. Apoptosis is very different from tissue necrosis caused by an acute injury, in which cells swell and burst (from osmotic pressure differences) and cause a significant immune response. Apoptosis consists of 4 steps:
The actual steps in cell death require:
To commit suicide must be an extremely important cellular decision. Hence you would expect this process to be regulated and highly complicated. When would it be advantageous to the organism to want a cell to kill itself (or be told to kill itself)? Cell death would be used to:
Unregulated apoptosis could exacerbate or cause disease such as:
Apoptosis does not require new transcription or translation, suggesting that the molecular machinery required for cell death lay dormant in the cell, and just requires appropriate activation. What "signals" induce apoptosis?
Signals can be extracelluar:
Signals can be intracelluar:
MECHANISMS AND REGULATION OF APOPTOSIS:
Caspases
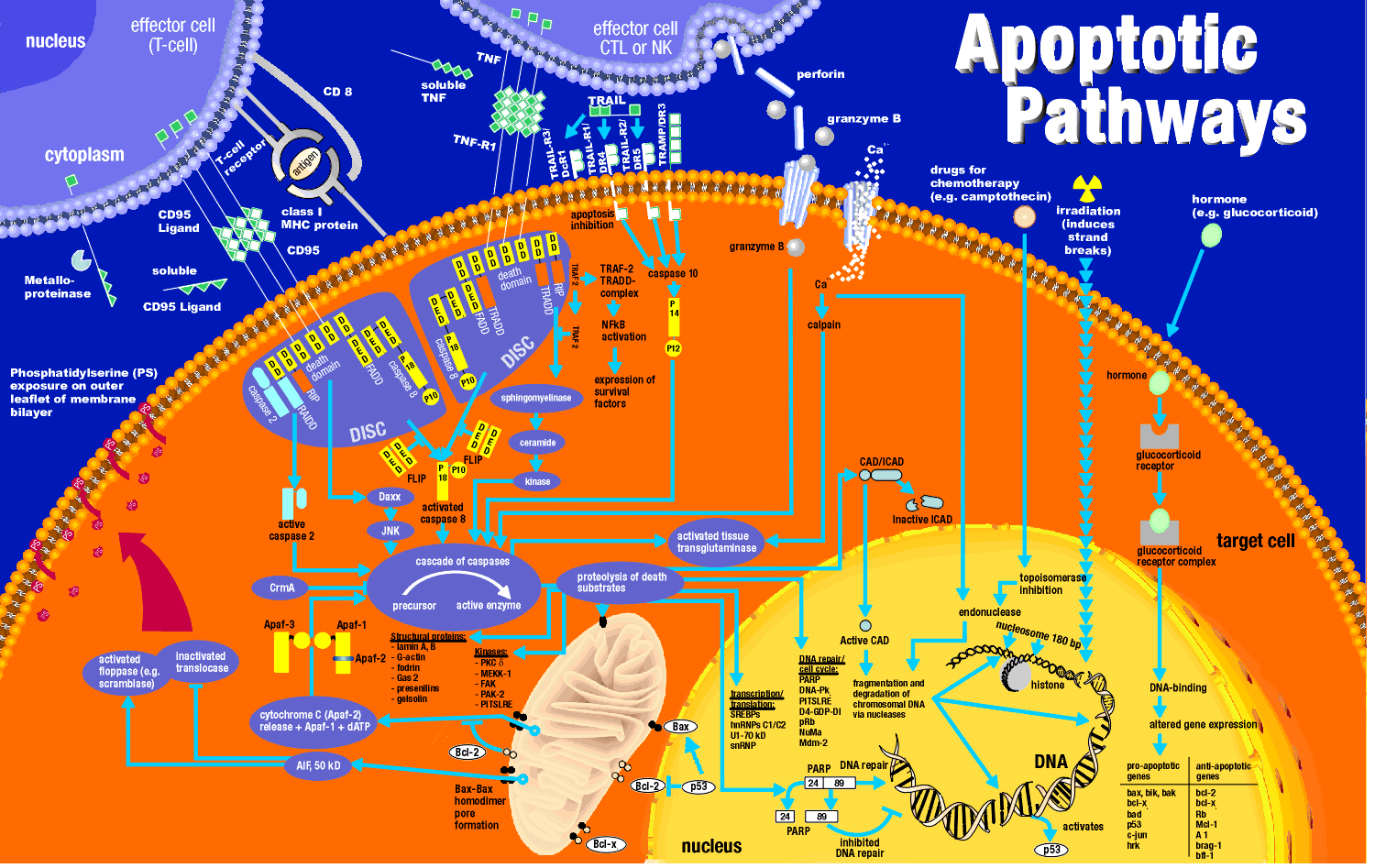
Characterization of apoptotic mechanisms and cellular players started with the study of C. elegans, a round worm. The mature worm has about 1000 cells. During development, 131 cells die. Two mutations were found in which the 131 cells did not die. These mutations were called ced3 and ced4 (ced stands for cell death). The sequence of ced 3 was very homologous to a protein called interleukin converting enzyme (ICE) which is required for proteolytic activation of the precursor to interleukin 1, a protein hormone released by certain immune cells during activation and which can promote inflammation. This suggested that proteolysis was required for apoptosis. Subsequent studies show that a whole family of proteases (about 10 in humans) called caspases (ICE has been renamed caspase 1) are required for programmed cell death. These proteases are found in the cell in an inactive form which must undergo limited proteolysis for activation. These caspases form a cascade of proteases which are activated in this process. They are endoproteases have an active site Cys (C) and cleave at the C-terminal side of Asp residues (asp) and hence are known as caspases - cys containing-asp specific proteases).
ICE is not normally involved in apoptosis, but its artificial activation in cultured mammalian cells can lead to it. Each caspase had the same sequence as they are designed to cleave, so it became evident that they probably cleave each other in an activation cascade mechanism, similar to the coagulation protease cascade of activation of precursors (zymogens) of serine proteases which activate the next in the series. Two series of caspases seem to be involved. One set initiates the process of caspase activation. Just as in the clotting system, the question of what activates the first caspase appeared problematic until investigators found that the initiator caspase can be activated if they aggregate to a critical concentration. This could occur by binding of a suicide signal molecule at the cell surface. Conformational changes in the receptor can lead to aggregation of surface receptor molecules with concomitant aggregation of intracellular caspases which interact with the aggregated receptors.
Intracellular signals
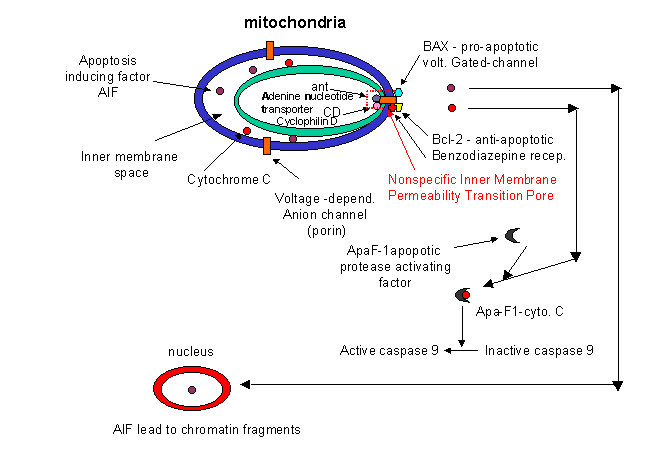
How might intracellular activators of apoptosis (like radiation or reactive oxygen species) work. Research indicated involvement of mitochondria in the apoptotic pathway. Believe it or not, cytochrome C, the heme protein which acts as a water soluble mobile carrier of electrons in mitochondrial oxidative phosphorylation, shuttling electrons through cytochrome C oxidase or complex IV, leaks out of the intermembrane space and binds to a cytoplasmic protein called Apaf-1 for apoptotic protease activating factor-1. This then activates an initiator caspase-9 in the cytoplasm.
These proteins seem to leak out of mitochondria after a collapse of the electrochemical potential across the inner membrane. The potential collapses as a consequence of the opening of a channel called a nonspecific inner membrane permeability transition pore, composed of both an inner membrane protein (adenine nucleotide translocator - ant) and an outer membrane proteins (porin, the voltage-gated anion channel - VDAC). These proteins act together, probably at sites where the inner and outer membranes are in contact. This channel passes anything smaller than molecular weight 1500. Collapsing the proton gradient uncouples oxidation and phosphorylation in the mitochondria. Changes in ionic strength causes a swelling of the matrix. Since the inner membrane is highly convoluted and has a much greater surface area than the outer membrane, swelling of the matrix leads to a rupture of the outer membrane, spilling the inner membrane space proteins (cytochrome C and Apaf-1) into the cytoplasm.
What causes all these changes in the mitochondria? Several interrelated events appear to be involved:
Caspase targets:
Apoptosis involves:
How does caspase activation lead to these events? A protein has been uncovered which when cleaved by a caspase leads to nuclear breakup. The target protein is usually bound to another protein, a DNA endonuclease. When the target protein is cleaved, the DNase is free to migrate to the nucleus and begin the execution. Membrane changes in apoptosis occur when caspase 3 cleaves gelsolin, a protein involved in maintaining cell morphology. The cleaved gelsolin cleaves actin filaments inside the cell. Another protein is necessary to form apopotic bodies: a kinase named p21-activated kinase 2 (PAK-2). This kinase is activated by caspase-3 by limited proteolysis. Caspases also cleave beta-amyloid precursor protein which might generate more beta-amyloid protein, causing neural cell death in Alzheimer patients.
Controlling Apoptosis
It should be clear that cells keep a tight control on the caspases. Two players which appear to inhibit apoptosis are the mitochondrial proteins Bcl-2 and Bcl-X, which can block the release of cytochrome C from the mitochondria. The Bcl family of proteins have a hydrophobic tail and bind to the outside surface of mitochondria and other organelles like the nucleus and endoplasmic reticulum. These proteins seem to be able to form ion channels in liposomes. So far 15 members of this family (related to ced-9 of C. elegans) have been discovered in humans. Bcl-2 can also bind to Apaf-1 (mentioned above) and inhibit its activation of initiator caspase-9. Bcl-2 is regulated by changes in the expression of the Bcl-2 gene, by post-translational phosphorylation by kinases, or by cleavage by caspases. Overexpression of Bcl-2 can cause a cell to become a tumor cell. Another member of the family, BAX and BAD bind to mitochondria and facilitate apoptosis by stimulating cytochrome C release.
In addition, other proteins called IAP's (inhibitors of apoptosis) can inhibit caspase or other apoptotic proteins. Some virus make the protease to keep their host cells viable.
Cell Membrane Events
Cells can be instructed to undergo apoptosis through cell surface interactions with other cells which are often immune cells. One of the jobs of the immune cell is to destroy an altered cell (for example a virally-infected cell or a tumor cell). Immune cells themselves must also die after they are activated in an immune response. Activated lymphocytes (like cytotoxic T cells or natural killer cells) can target and kill cells using several ways which can involved apoptosis. In one, an activated lymphocyte binds to a target cell (like a virally infected cell) and secrete perforin, a protein which assemble in the target cell membrane to form a transmembrane channel. Other proteins released by the activated lymphocyte can enter the target cell through the pore and initiate apoptosis. One such protein that enters, granzyme B is a protease which activates caspases in the target cell.
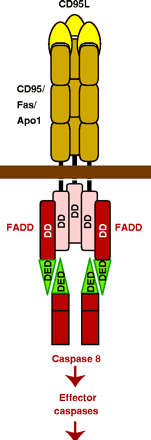
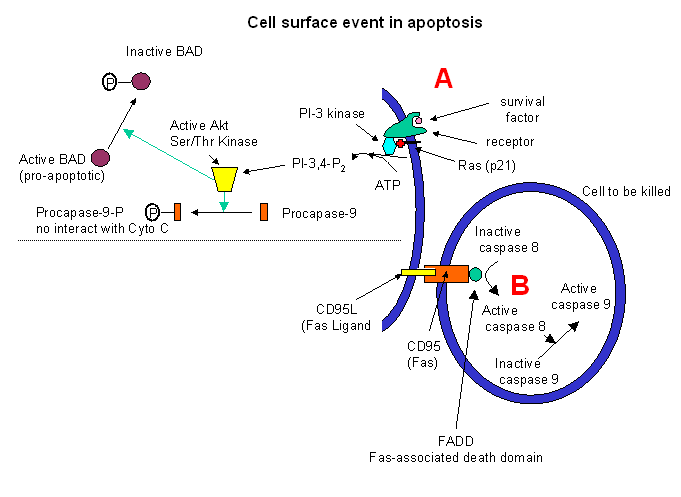
Target cells which express a specific membrane protein call CD95 (also called Fas) are also targeted for apoptosis. This protein receptor, a member of the tumor necrosis factor receptor (TNFR) binds to a membrane protein ligand on the surface of an activated lympocyte called CD95 Ligand - CD95L- (also called the Fas ligand). On binding, the CD95 (Fas) receptors on the target membrane aggregate after conformation changes. An adapter protein in the cell, FADD (Fas-associated death domain) binds to the aggregated cytoplasmic domain (the death domain) of CD95 (Fas), and recruits inactive caspase-8 to the site, where their concentration increases. This leads to activation of the caspases.
This mechanism is used to get rid of activated lymphocytes after they have finished their work. Activated immune cells start expressing Fas a few days after activation, targeting them for elimination. Some cells which have been stressed express both Fas and Fas ligand and kill themselves. Various cells express CD95 (Fas), but CD95L (Fas-Ligand) is expressed predominately by activated T cells.
Cell surface events also can inhibit apoptosis. Binding of "survival" factors (like growth factors) to cell surface receptors can shut of apoptotic pathways in the cells. Some survival factor receptors are coupled to PI-3-kinase (phosphoinositol-3-kinase) through the G protein ras (p21) which is targeted to the cell membrane by post-translational addition of a hydrophobic anchor. The activated kinase produces PI-3,4-P2 and PI-3,4,5-P3, which activates Akt, a Ser/Thr protein kinase. This activated kinase phosphorylates the proapoptotic-protein BAD, which then becomes inactive. In addtion, active Akt phosphorylates procapse, which in its phosphorylated form will not interact with cytochrome C, hence inhibiting apoptosis.
The endpoint of apoptosis is the engulfment of the fragmented cell by a phagocytic cell (such as a macrophage). In a recent article (Nature, 405, pg 85, 2000), it was shown that the activity of phagocytes could be inhibited sterospecifically by the addition of phosphatidyl serine (PS) to the mixture, but not by other negative phospolipids. If you remember from our description of lipids, PS is found exclusively in the inner leaftlet of red blood cells). The investigators cloned a gene from the phagocytic cell for a receptor which recognizes PS. When added to ordinary T and B lymphocytes (immune cells), these cells could also take-up apoptotic cells. The gene is homologous to genes in Drosophila (fruit fly) and C. elegans (round worm) suggesting that it is conserved in nature. The message: when cells undergo apopotosis, PS, normally found only in the inner leaftlet, is exposed to the outside. It can then bind to receptors on phagocytic cells to complete the process of apoptosis.
Web Sites for Apoptosis:
References:
Navigation
{kind=link}
{kind=link}
{kind=link}
{kind=link}