Biochemistry Online: An Approach Based on Chemical Logic

CHAPTER 1 - LIPID STRUCTURE
C: Dynamics of Membrane Lipids
BIOCHEMISTRY - DR. JAKUBOWSKI
02/12/2016
Learning Goals/Objectives for Chapter 1C: After class and this reading, students will be able to
|
C1. Dynamics of Membranes
Molecules are not static, but rather are dynamic. This also applies to molecular aggregates. In the first part of the section, we will discuss the rigid movement of whole lipid molecules in a bilayer, within a leaftlet and between leaflets. In the second part and the following supplement, we will consider the movement of atoms within a molecule. The movements include motions like bond bending, bond stretching and torsion angle changes like we saw in the previous chapter section on the conformations of n-butane. The position of all atoms within a molecule can be simulated as a function of time - a molecular dynamics simulation. Such motions affect the energy of the molecule, which can be calculated for given atom positions using classical molecular mechanics and electrostatics.
Liposomes and bilayers in general must be somewhat dynamics, otherwise they would be impenetrable barriers across which nothing could pass. Cell membranes must separate the outside of a cell from the inside, but they must also allow passage of molecules and even ions across the membrane. What is the evidence that membranes are dynamic?
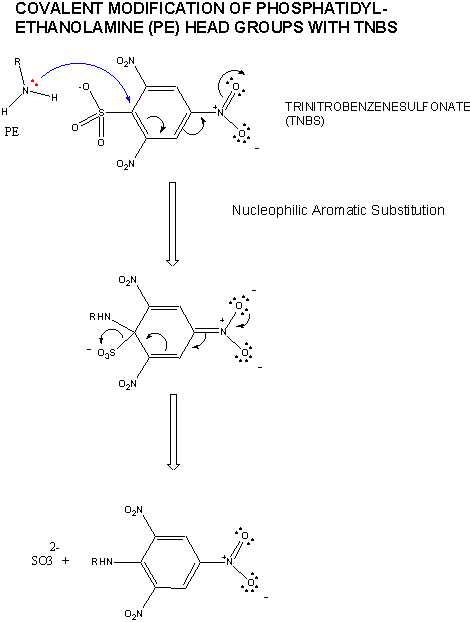
First, lipids can diffuse laterally in the membrane. This can be shown as follows. Make a liposome from phosphatidylethanolamine, PE, which has been labeled with TNBS (Trinitrobenzensulfonate). The NH2 on the head group of PE can attach the TNBS which undergoes nucleophilic aromatic substitution with the expulsion of the SO3-2. The TNB group attached to the PE head group absorbs UV light and emits light of higher wavelength in a process called fluorescence. Next, using a fluorescent microscope, the fluorescence intensity of a region of the surface can be recorded. Then shine a laser on a small area of the surface, which can photobleach the fluorescence in the area. Over time, fluorescence can be detected from the region again. The rate at which it returns is a measure of the lateral diffusion of the labeled lipids into the region. Lipids can undergo lateral diffusion at a rate of about 2 mm/s. This implies that the lipids can transit the surface of a bacteria in 1 sec.
{kind=link}
{kind=link}
Transverse, or flip-flop diffusion (movement of a phospholipid from one leaftlet to the other, not within a given leaflet) should be more difficult. Experimentally, this is investigated as shown in the diagrams below.
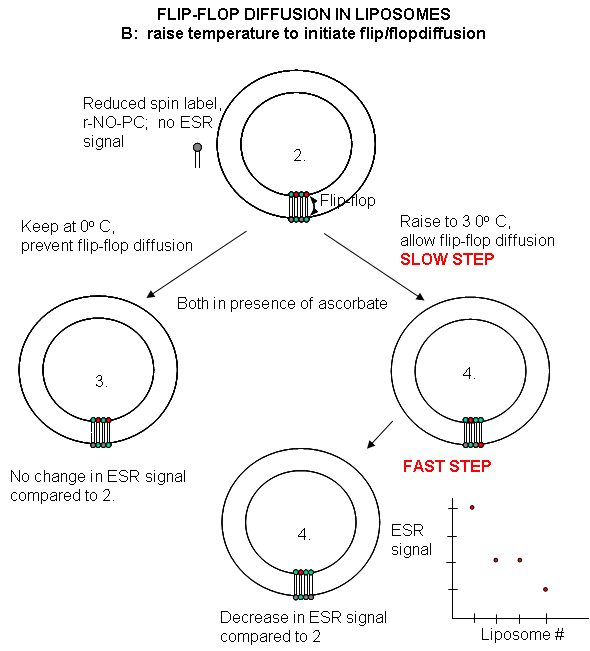
Liposomes experiments: To test flip-flop diffusion in an artificial membrane, liposomes are made with a mixture of PC and a PC derivative with a nitroxide spin label (has a single unpaired electron). Both inner and outer leaflets of the membrane have the labeled PC. Like a proton in NMR spectroscopy, a single electron has a spin which can give rise to an electron-spin resonance (ESR) signal (as a proton gives rise to a nuclear magnetic resonance signal) when irradiated with the appropriate frequency electromagnetic radiation (microwave frequency for ESR, radio frequency for NMR) in the presence of a magnetic field. The liposomes are kept at 0oC and the ESR signal is determined. Ascorbic acid, a water soluble vitamin and antioxidant/ reducing agent, is added to the liposomes. This reduces the spin labeled PC in the outer leaflet, but not the inner leaftlet of the bilayer since ascorbic acid can not enter the liposome or otherwise interact with it. This reduces the ESR signal to a lower, constant value. The sample is divided into two. One sample is left at 0oC, the other is raised to 30oC. The ESR signal is recorded as a function of time. The 0oC prep shows no change in ESR with time, while the 30oC prep ESR signal decreases with time. This decrease resultsfrom flip-flop diffusion of labeled PC from the inner leaflet to the outer, and subsequent reduction by ascorbic acid. These experiments in experimental bilayer systems like liposomes shows that flip-flop diffusion is orders of magnitude slower than lateral diffusion.
Figure: Flip/Flop diffusion in liposomes A: Making vesicles with ESR active PC analog only in outer leaflet
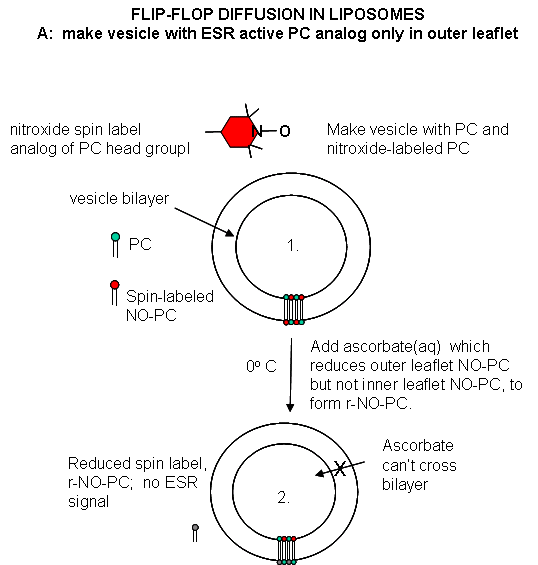
Figure: Flip/Flop diffusion in liposomes B: Raising the temperature to initiate flip/flopdiffusion
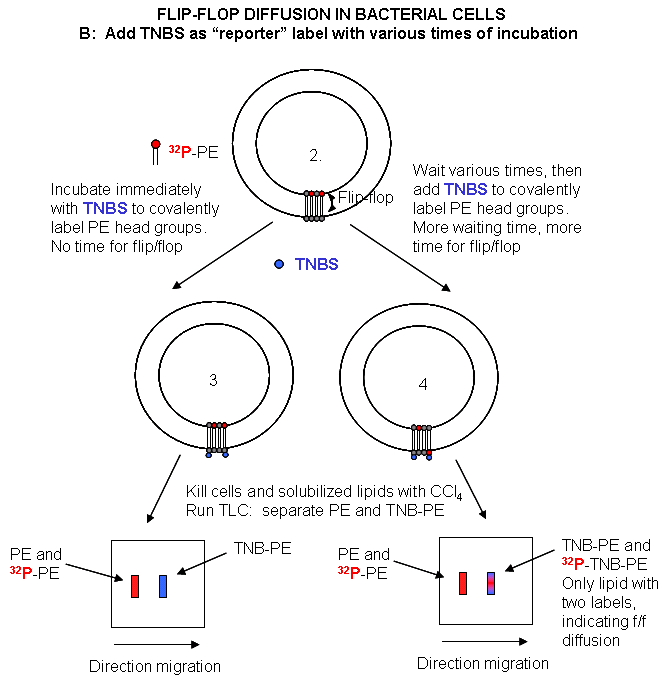
Cell Experiments: An analogous experiment can be done with bacteria. Radiolabeled 32 PO4 is added to cells for one minute, which leads to the labeling of newly synthesized phospholipid (PL) which locates to the inner leaflet. The cells are then split into two samples. One sample is reacted immediately with TNBS, which will label only PE in the outer leaflet. The other sample is incubated 3 minutes (to allow PL synthesis) and then reacted with TNBS. After a short labeling period, the cells are destroyed by adding organic solvents which prevents new lipids biosynthesis. The lipids are extracted into the solvent and then subjected to TLC.
The lipids can be labeled in three ways. Some will be labeled with 32 P alone, some with TNBS alone, and some with both 32P and TNBS. TLC (or other techniques such as HPLC or GC) can easily separate PC and TNBS-labeled PC since they have different structures and hence will migrate to different places on a TLC plate. No chromatographic technique could, however, separate PC and 32 P-PC, since their molecular structure is the same, the only difference being in the nuclei of the P (different number of neutrons).
Those lipids with double labels (TNB and 32 P) must have flipped from the inner leaftlet to the outer leaftlet where they could be labeled with TNBS. The cells incubated for 3 minutes before the addition of TNBS have a much higher level of doubly labeled PL's. Quantitating these data as a function of differing time of incubation at elevated temperatures show that the rate of flip-flop diffusion is much higher in cells than liposomes, which suggests that the process is catalyzed, presumably by a protein transporter (flipase or Transbilayer amphipath transporter - TAT) in cells.
Figure: Flip/Flop diffusion in bacterial cells A: Labeling inner leaflet phospholipids with 32 P
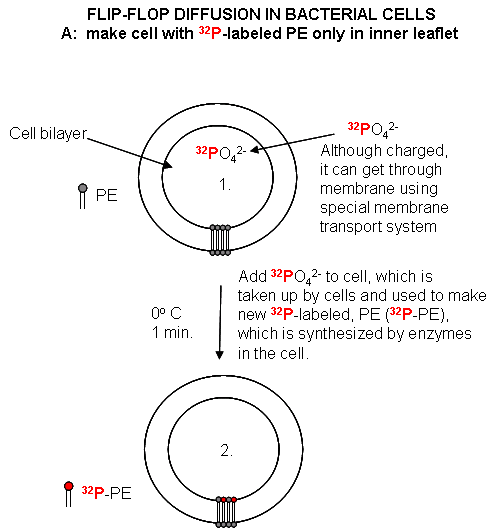
Figure: Flip/Flop diffusion in bacterial cells B: Labeling Cells in A with TNBS to detect Flip/Flop
 Navigation
Navigation
Return to Chapter 1C: Dynamics of Membrane Lipids
Return to Biochemistry Online Table of Contents
Archived version of full Chapter 1C: Dynamics of Membrane Lipids