Biochemistry Online: An Approach Based on Chemical Logic

CHAPTER 9 - SIGNAL TRANSDUCTION
E: Metabolic Control Analysis and
Systems Biology
BIOCHEMISTRY - DR. JAKUBOWSKI
04/16/16
|
Learning Goals/Objectives for Chapter 9E:
|
E3. Metabolic Control Analysis and Simple Enzyme Inhibition
Biochemists model complex enzyme-catalyzed reactions in the presence
and absence of modifiers (either activators or inhibitors) to develop
mechanisms for the reactions. The following kinetic parameters are
experimentally determined by fitting initial velocity (vo) vs substrate
concentration ([S]) through nonlinear fitting algorithms.
- Km – the Michaelis Constant, the concentration of substrate at half-maximal velocity;
- Vm – the velocity at saturating substrate concentration;
- kcat - the turnover number for conversion of bound reactant to product;
- Kix – inhibition dissociation constants.
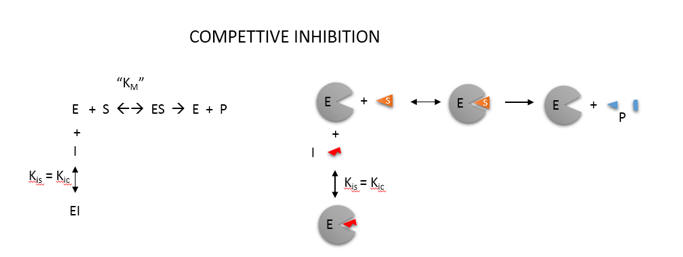
An example of competitive inhibition illustrates a common type of
analysis for such reactions.
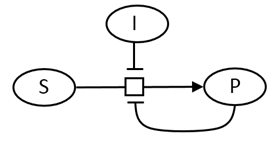
A modern pictorial depiction of a simple irreversible, enzyme-catalyzed reaction of substrate S going to product P with inhibition by the product and by an added inhibitor is shown below:
Consider the simple enzyme-catalyzed reaction for a reversible conversion of substrate S to product P that has 3 reversible steps.
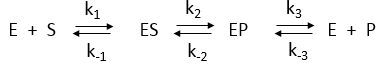
If the forward (f) and reverse (r) chemical reaction steps (reactions 2 and -2) were irreversible and written separately, simple Michaelis-Menten equations could be written for each
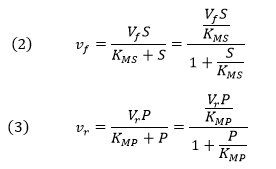
For the actual reversible reactions for step 2, the net forward rate cannot be found by simple subtraction of the two equations above as the differential equations describing the simple forward and reverse rates don’t account for the reverse steps
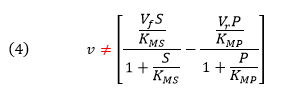
A simple derivation (assuming rapid equilibrium for both forward and reverse steps) can be made for the net forward reaction. Again consider the following enzyme catalyzed reaction:
You may remember that for the isolated E + S <----> ES and for E + P <----> EP reactions, the simple dissociation constants, KS and KP are given by
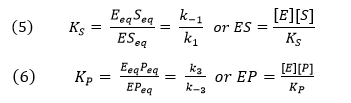
The rapid equilibrium assumption states that the rate of dissociation of
ES and EP, which are both physical steps, are much faster than the rate of
the chemical conversion steps for each complex. Hence k-1 >> k2 and k3 >>
k-2, so the relative amounts of ES and EP can be determined from the
dissociation constants as shown above.
Mass conservation of enzyme gives
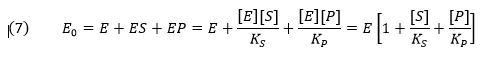
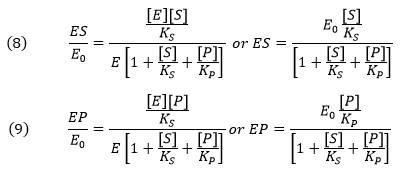
From this we can get the fractional amount of both ES and EP
Now we can derive the rate equation
for the net forward reaction for the rapid equilibrium case:
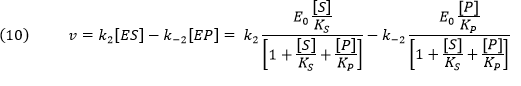
Knowing that k2E0 and k-2E0 represent the maximal velocities, Vf and Vr, respectively, the equation becomes:
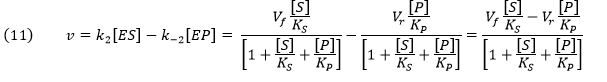
An equation of similar form can be derived from the steady state assumption. This equation is clearly different from the earlier equation (4) derived by intuitively assuming a simple subtraction of the irreversible forward and reverse rates which we have now shown to be invalid by comparison.
Programs like COPASI have many built in equations for velocities of many enzyme-catalyzed reactions that have similar forms. Two are shown below:
Reversible Michaelis-Menten: 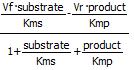
Competitive Inhibition Reversible:
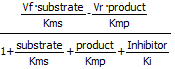
 Navigation
Navigation
Return to Chapter 9E: Metabolic Control Analysis and Systems Biology Sections
Return to Biochemistry Online Table of Contents

Biochemistry Online by Henry Jakubowski is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.