Biochemistry Online: An Approach Based on Chemical Logic

CHAPTER 9 - SIGNAL TRANSDUCTION
B: NEURAL SIGNALING
BIOCHEMISTRY - DR. JAKUBOWSKI
06/10/14
|
Learning Goals/Objectives for Chapter 9B:
|
B2. Transmembrane Potentials
Several questions arise about the distribution of ions and the magnitude of the transmembrane potential.
- How are the ion gradients established?
- How does the transmembrane ion distribution contribute to the membrane potential?
- How can the resting electrochemical potential and the ion distribution be maintained?
The answer to these questions will be
illustrated using studies on two types of brain cells, glial cells (which
function as protectors, scavengers, and feeder for brain neurons) and
neurons. Both types of cells have transmembrane potentials.
Glial Cells
1. The transmembrane ion gradients for ions can
be established by different mechanisms. One uses ion-specific ATPases
(P-type ion transporters), such as we discussed with the Na/K ATPase. This
transporter ejects 3 sodium ions from the inside of the cell for every 2
potassium ions it transports in, all against a concentration gradient.
Since it is an electrogenic antiporter, it helps generate the potential.
Additionally, specific ion channels also contribute (as described below) to
the transmembrane gradients and potentials.
2. The harder question
is how the ion distribution contribute to the membrane potential. Two
things must occur for a membrane potential to exist: First, there must
be a concentration gradient of charged ions (for example, sodium, potassium,
or chloride) across the membrane. Second, the membrane must be
differentially permeable to different ions. If the membrane were
completely impermeable to ions, then no movement of ions across the membrane
could occur, and no membrane potential would arise. If, however,
membranes are differentially permeable to the ions, an electrical potential
across the membrane can arise. Remember, synthetic bilayers are
quite impermeable to ions, given the hydrophobicity of the internal part of
the bilayer. Likewise it is quite impermeable to glucose. It turn out that
glial cells appear to have only a non-gated potassium channel, which allows
the outward flow of potassium ions down the concentration gradient. The
inside will then have a net negative charge since impermeable anions remain.
The chemical potential gradient causes this outward flow of potassium ions.
As more ions leave, the inside gets more negative, and a transmembrane
potential develops which resists further efflux of potassium. Eventually
they balance, and the net efflux of potassium stops. The resting
transmembrane potential reaches -75 mV which is exactly the value obtained
from the equations we will derive below. Since glial cells appear to only
express a nongated potassium channel, their resting potential is equal to
the potassium equilibrium potential.
Figure: Visualizing the transmembrane potential in K+ loaded vesicles + a nongated K+ channel.
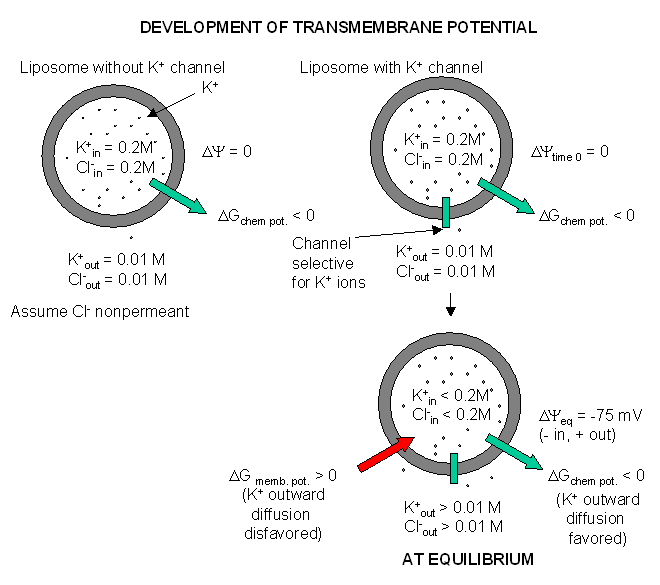
(Note: above figure has mistake. Cl- ion , assuming that is not permeable, will not change across the membrane at equilibrium.)
 Navigation
Navigation
Return to Chapter 9B: Neural Signaling Sections
Return to Biochemistry Online Table of Contents
Archived version of full Chapter 9B: Neural Signaling

Biochemistry Online by Henry Jakubowski is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.